Embriologia
Tratto dall'opera online "APUNTES DE EMBRIOLOGIA HUMANA" del Profesor Jano
Traduzione e adattamento di Giorgio Beltrammi
L'embriologia è la branca della biologia che studia i processi tramite i quali gli organismi crescono e si sviluppano prima della nascita.
L'embriologia odierna studia tutte quelle tappe che da un gamete femminile (ovulo o cellula uovo) fecondato da un gamete maschile (spermatozoo) portano ad un individuo maturo adulto. Il termine embriologia, perciò si riferisce allo studio degli organismi tra lo stadio unicellulare (generalmente, lo zigote) e l'inizio della vita indipendente. [Wikipedia]
1. Gametogenesi
- Origine extraembrionaria delle gonadi (tappa comune per maschio e femmina)
Le prime cellule germinali cominciano a formarsi a partire dal 24mo giorno di gestazione nel foglietto endodermico del sacco vitellino.
Da li migrano ai primordi gonadici che sono le sedi nelle quali si stanno formando le gonadi. A volte queste cellule si dislocano e vanno in altre sedi. Normalmente muoiono, ma se proliferano generano teratomi. I teratomi sono masse tumorali rigonfie che contengono diversi tipi di tessuti differenziati come pelle, peli, cartilagine e inclusi denti. Le sedi più frequenti nelle quali si possono incontrare questi tumori sono la bocca e la regione sacrococcigea.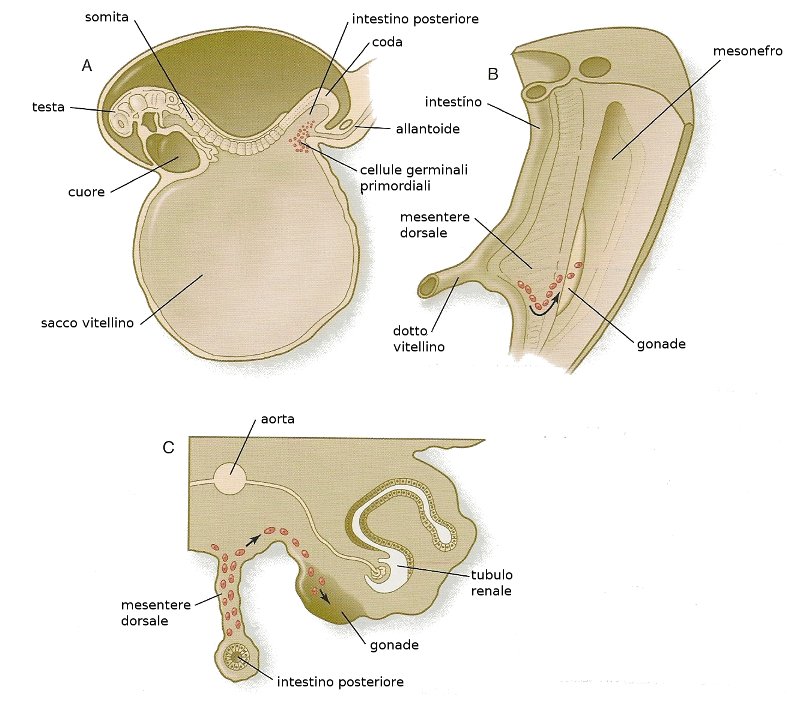
- Proliferazione delle cellule germinali
Quando le cellule germinali vanno ai primordi germinali si dividono molte volte per mitosi, generando cellule uguali e tutte diploidi. Queste cellule sono:
- Spermatogoni:
- Si formano per tutta la vita del maschio
- La proliferazione di queste cellule è ad intervalli
- Alloggiano nei tubi seminíferi del testícolo
- queste cellule, per meiosi e maturazione, origineranno gli spermatozoi
- Ovogoni:
- Si producono tra il secondo ed il quinto mese di gravidanza generando tutta la popolazione degli oogoni che avrà una femmina per tutta la sua vita: 7.000.000 come massimo. Solo 2.000.000 arriveranno a maturazione.
- Per un processo di atresia (degenerazione naturale) molti oogoni vanno van degenerando lungo la vita della femmina fino alla menopausa.
- Spermatogoni:
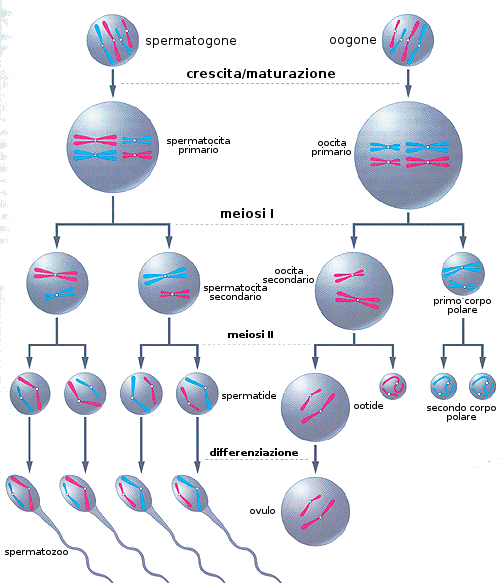
- Produzione dei gameti
In questa tappa si produce la riduzione a metà del numero dei cromosomi della specie per permettere nuovamente la condizione diploide dopo la fecondazione.
- Nel caso della femmina:
Si inizia per meiosi che comincia nel periodo fetale. Gli oogoni che entrano in meiosi ricevono il nome di oociti primari, nome che mantengono fino al termine della I divisione meiotica. Dopo la nascita non si formano più oociti primari. La caratteristica principale di questa meiosi femminile sono i suoi blocchi.
- Blocco I: si produce nel diploteno della profase I. Rimangono così le cellule fino alla pubertà. Questo è lo stato dell'Ovocito primario che perdura fino alla pubertà. Una volta passato questo periodo della vita, ogni 28 giorni tra i 10 ed i 30 ovociti primari completano la I divisione meiotica poco prima della ovulazione producendo due cellule figlie: l'ovocito secondario e il primo corpo polare. Continua la II divisione meiotica fino al:
- Blocco II: si produce nella metafase II. La cellula terminerà la meiosi si è stata fecondata da uno spermatozoo.
- Nel caso del maschio
La meiosi degli spermatociti primari dei tubi seminiferi del testicolo comincia nella pubertà. A partire da essa e in modo ininterrotto, si producono spermatozoi. Dopo 24 giorni e attraverso la divisione meiotica I si formano gli spermatociti secondari (con n cromosomi) che inizieranno la divisione meiotica II che dopo otto ore produrranno quattro spermatidi
- Nel caso della femmina:
Si inizia per meiosi che comincia nel periodo fetale. Gli oogoni che entrano in meiosi ricevono il nome di oociti primari, nome che mantengono fino al termine della I divisione meiotica. Dopo la nascita non si formano più oociti primari. La caratteristica principale di questa meiosi femminile sono i suoi blocchi.
- Maturazione dei gameti
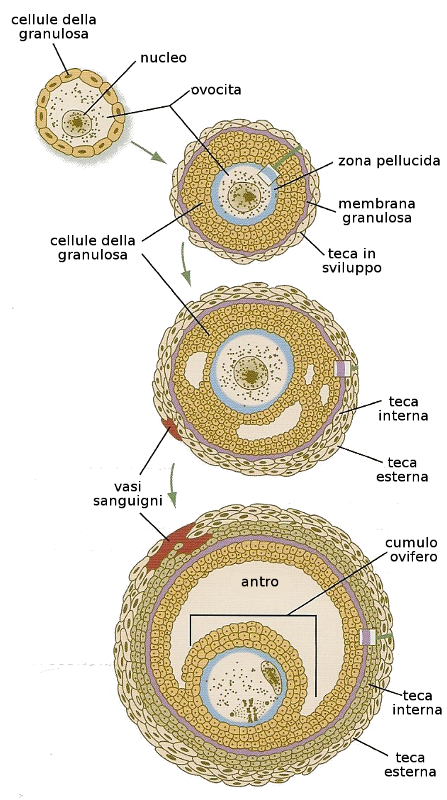 Nel caso degli oociti: l'ovocita primario nello stato fetale si contorna di uno strato di cellule di tessuto connettivo costituendo i follicoli primordiali. Nella pubertà, insieme all'aumento della grandezza dell'ovocita primario, le cellule follicolari acquisiscono forma cilindrica e si costituisce il follicolo primario. Più avanti questo strato di cellule prolifera e passano a costituire vari strati e successivamente si contornano di una coperta di materiale glicoproteico che riceve il nome di zona pellucida. Queste cellule che accompagnano l'ovocita secondario e che formano la maggior parte del follicolo, si chiamano cellule della granulosa.
Nel caso degli oociti: l'ovocita primario nello stato fetale si contorna di uno strato di cellule di tessuto connettivo costituendo i follicoli primordiali. Nella pubertà, insieme all'aumento della grandezza dell'ovocita primario, le cellule follicolari acquisiscono forma cilindrica e si costituisce il follicolo primario. Più avanti questo strato di cellule prolifera e passano a costituire vari strati e successivamente si contornano di una coperta di materiale glicoproteico che riceve il nome di zona pellucida. Queste cellule che accompagnano l'ovocita secondario e che formano la maggior parte del follicolo, si chiamano cellule della granulosa.
Quando giunge la pubertà si aggiunge al follicolo un doppio strato di cellule del proprio ovaio che costituiscono le teche (interna ed esterna) che tra le altre sostanze secernono fattori di angiogenesi. Inoltre, con la secrezione del FHS (ormone follicolo stimolante) le ovaie cominciano a produrre estrogeni e alcuni follicoli accumulano liquido in una cavità che si chiama antro. In questo stato il follicolo passa a chiamarsi follicolo secondario.
Alcuni follicoli secondari aumentano di dimensione per crescita dell'antro e passano a chiamarsi follicoli di Graaf o terziari. Un ovaio di una femmina fertile contiene i tre tipi di follicoli. Le cellule del follicolo sono quelle che secernono un fattore inibitore della meiosi che fa si che questo processo rimanga bloccato nel diplotene (primo blocco)- Nel caso degli spermatociti: Si è già visto che gli spermatociti entrano in meiosi nei tubi seminiferi. Insieme a queste cellule si trovano le cellule di Sertoli che sostengono e nutrono gli spermatociti. Quando questi giungono allo stato di leptotene si staccano dalla barriera che formano le cellule di Sertoli ed salgono dall'interno dei tubi seminíferi. Li completeranno la I Divisione Meiotica trasformandosi in spermatociti secondari. Otto ore più tardi hanno già completato la Divisione Meiotica II lasciando quattro spermatidi per ogni spermatogonia di tipo B che è entrata in meiosi.
Mediante il processo di spermiogenesi gli spermatidi maturano trasformandosi in spermatozoi. I principali cambi prodotti sono:- riduzione del nucleo e gran condensazione della sua cromatina
- Formazione dell'acrosoma (una gran vescicola piena di enzimi idrolitici)
- Generazione del flagello
- Si accumulano mitocondri nella regione prossimale del flagello.
Tutto il processo di formazione di uno spermatozoo dura 64 giorni. Questa cellula aploide è molto specializzata e ben adattata al movimento. Consta di una testa di 2-3 micrometri di larghezza e 4-5 micrometri di lunghezza, che contiene il nucleo e l'acrosoma. La zona con la quale la testa si congiunge al flagello si chiama zona intermedia e contiene la regione prossimale del flagello contornato da una spirale di mitocondri. Per ultima si trova la coda, o flagello che ha una lunghezza di circa 50 micrometri.
Giorgio Beltrammi
Bio-Pedia III
648 pagine B/N, 1683 termini circa
Formato A5 (148 x 210 mm.)
Acquista →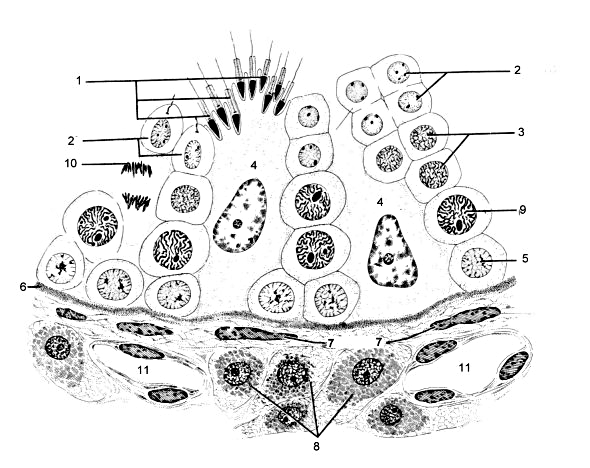
1. Spermatozoo; 2. Spermatide; 3. Spermatocita secondario; 4. Cellula di Sertoli; 5. Spermatogonia; 6. Lamina basale; 7. Fibroblasto; 8. Cellule interstiziali o di Leydig; 9. Spermatocita primario; 10. Cellula in meiosi; 11. Vaso sanguigno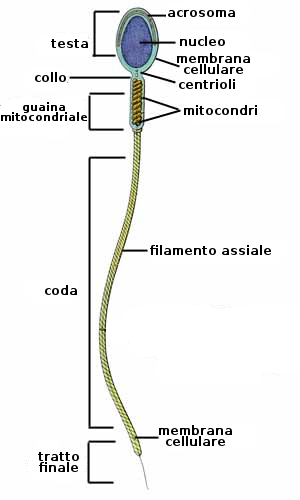
 |   |
2. Fecondazione
2.1 Ovulazione
Verso la metà del ciclo mestruale (in teoria il 14mo giorno dei 28) un follicolo di Graaf maturo che contiene l'ovulo bloccato in diplotene, si approssima alla superficie dell'ovaio e aumenta di grandezza per azione dell'ormone follicolo stimolante (FSH) e dell'ormone luteinizzante (LH). Sotto questo stimolo completa la I divisione meiotica e viene espulso il primo corpo polare. In questo stato il follicolo va verso la superficie dell'ovaio. Dopo 28 - 36 ore dal picco dell'LH avviene la rottura del follicolo e l'espulsione dell'ovulo dall'ovaio verso la cavità peritoneale. È l'ovulazione.
L'ovulo viene espulso con degli involucri accompagnatori:- La zona pellucida
- Uno strato di due o tre cellule che forma la corona radiata
- Uno strato di sostanza appiccicosa

2.2 Eiaculazione e trasporto degli spermatozoi
Dopo la spermiogenesi gli spermatozoi si accumulano nell'epididimo dove acquisiscono la maturità biochimica ad essere capaci di fecondare l'ovulo. In questo luogo passano circa dodici giorni. Nella eiaculazione gli spermatozoi si riversano nel dotto deferente e si mescolano con la secrezione delle vescicole seminali e della prostata. Le prime apportano un 60% del seme con una secrezione a carattere basico con abbondanza di fruttosio come fonte nutritiva per gli spermatozoi, e la seconda il 30 % con una secrezione acida ricca di acido citrico, zinco e magnesio.
Nella copula, il seme si deposita solitamente nel fornice anteriore della vagina che ha una secrezione acida ad azione battericida. Il seme a carattere alcalino protegge gli spermatozoi da questo ambiente acido ostile, dando loro il tempo di guadagnare il collo uterino dove incontrano un ambiente ottimo per la loro
motilità con un pH tra 6 e 6,5.
2.3 Il muco cervicale
La composizione e la viscosità delle secrezioni mucose de collo dell'utero variano molto durante il ciclo mestruale.- Tra i giorni 9 e 16 del ciclo aumenta il contenuto in acqua per facilitare il passaggio degli spermatozoi. Riceve il nome di muco E.
- Dopo l'ovulazione e per effetto del progesterone si passa ad un muco più viscoso per una minor proporzione di acqua. È il muco G che è praticamente resistente alla penetrazione degli spermatozoi.
2.4 Fecondazione
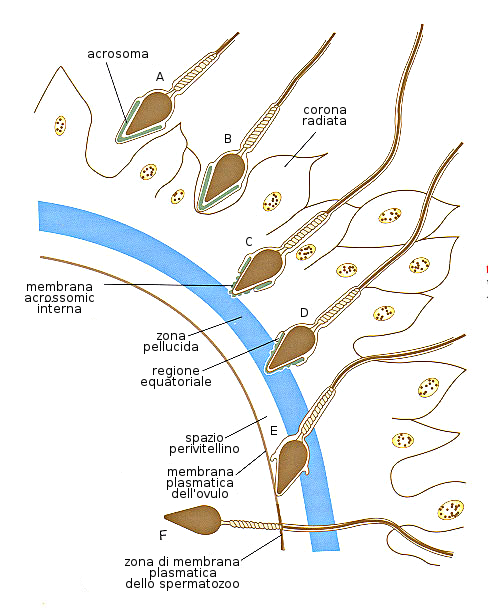
La fecondazione solitamente avviene nel terzo superiore della Tuba di Fallopio che ha captato l'ovulo. Comincia con la penetrazione dello spermatozoo nella corona radiata grazie al movimento del flagello e all'azione della ialuronidasi e altri enzimi dell'acrosoma. Attraversa anche i seguenti strati: la membrana plasmatica dell'ovocita e la zona pellucida. Dopo la fusione della membrana plasmatica dello spermatozoo con quella dell'ovocita, il gamete maschile penetra nel citoplasma dell'ovocita e questo conclude la II divisione meiotica formandosi l'ovocita maturo e il secondo corpo polare.
La coda dello spermatozoo degenera e fonde il suo nucleo con quello dell'ovocita: si forma lo zigote e si ricupera la condizione diploide (2n)
- Uno rapido che consiste in un cambio del potenziale della membrana a 2-3 secondi dalla fecondazione e dura cinque minuti.
- Uno lento che presuppone la fusione dei granuli corticali prossimi alla membrana interna all'ovocita, per formare uno strato isolante che impedisce la penetrazione di più spermatozoi.
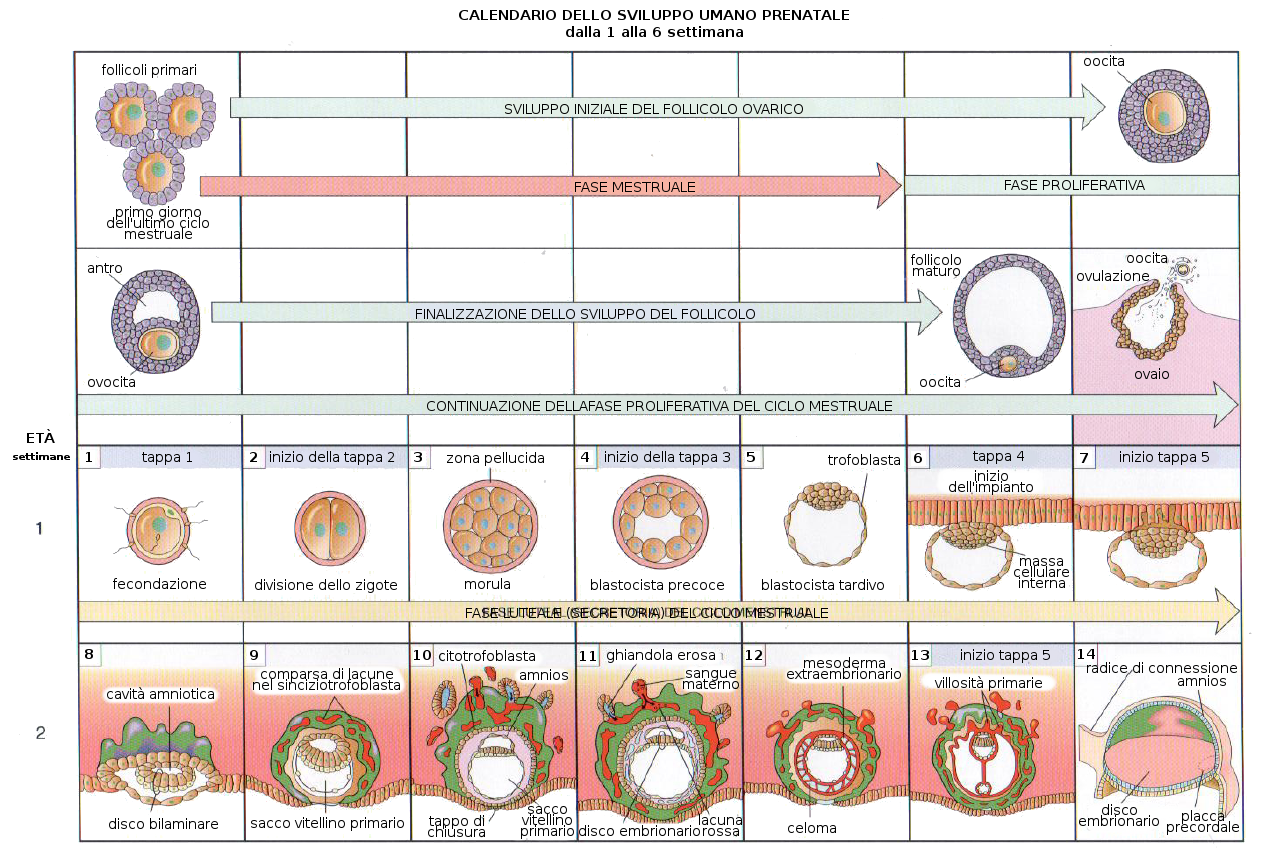
3. Segmentazione e Impianto
Dopo la fecondazione, comincia la prima tappa dello sviluppo embrionario: la segmentazione. Questa consiste in una serie di mitosi che producono una serie di cellule embrionarie denominate blastomeri. Dopo tre giorni di divisioni si giunge ad un stato di 16 cellule compatte che riceve il nome di morula.
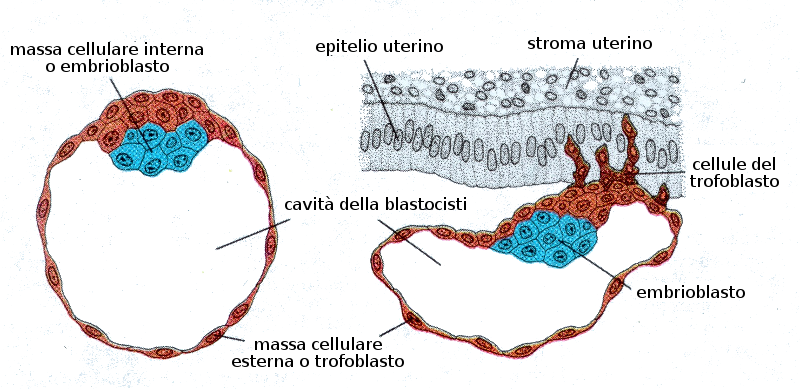
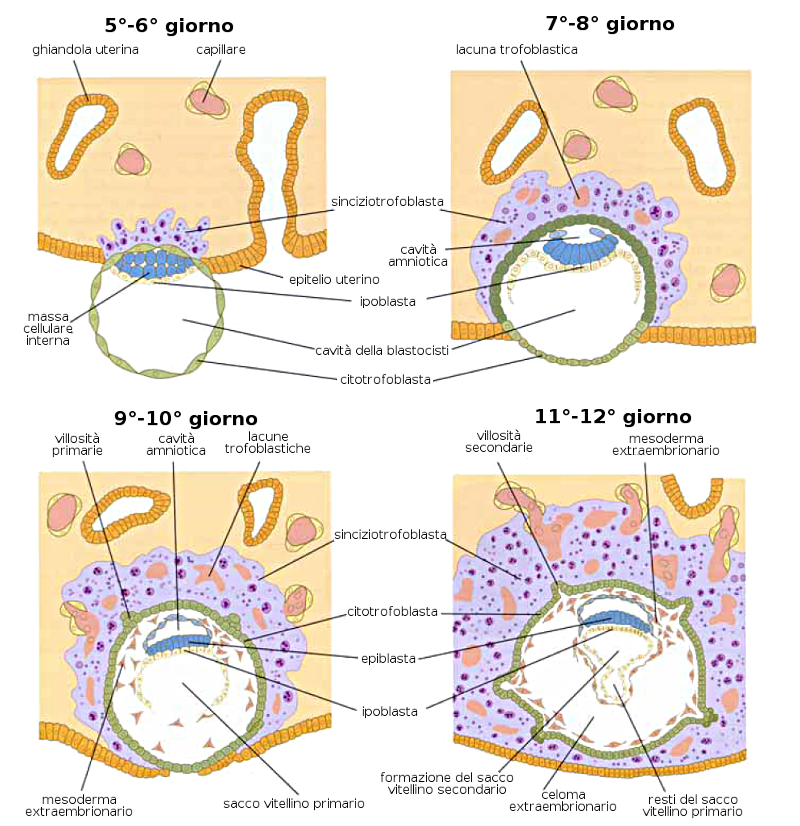
Al quarto giorno di gestazione e dopo un processo nel quale interviene la pompa del Na+ e K+ dei blastomeri si produce una entrata di acqua all'interno della morula che genera una cavità chiamata blastocele (o cavità del blastocisto). La massa cellulare interna costituisce l'embrioblasto. Questo stadio si conosce con il nome di blastocisti. È il quinto giorno dopo la fecondazione.
- Uno strato esterno di cellule che produrrà il trofoblasto che a sua volta darà la placenta, e
- Una massa cellulare interna che sarà l'embrione propriamente detto o embrioblasto. Questa regione riceve il nome di polo embrionario e quella opposta abembrionario.
Questa prima differenziazione è condizionata dal contatto tra cellule: per differenziarsi in cellule del trofoblasto queste devono essere in contatto con superfici non adesive come quelle della zona pellucida che le contorna. questa zona pellucida scompare poco prima dell'impianto, ma le cellule della periferia sono già state "marcate" per differenziarsi in trofoblasto.
Allo stato di blastocisti e sei giorni dopo la fecondazione avviene l'Impianto dell'embrione nella mucosa uterina (endometrio) della madre nella regione del polo embrionario. In concreto, le cellule del trofoblasto sono quelle che si introducono nella mucosa grazie alla produzione di enzimi proteolitici che si fanno largo nell'endometrio. Questo succede verso il sesto giorno di gravidanza.

3.1 Disco germinativo bilaminare
A partire dall'ottavo giorno l'embrione è completamente impiantato nell'utero della madre e le cellule sono totipotenti e senza differenziazioni. Ora comincia la prima differenziazione e le due parti della blastocisti si suddivide in:- Trofoblasta:
- Citotrofoblasta: strato interno di cellule mononucleate
- Sinciziotrofoblasta: zona esterna di cellule multinucleate
- Embrioblasta:
- Ipoblasta: strato di cellule cubiche adiacente alla cavità del blastocista
- Epiblasta: strato di cellule alte adiacenti alla cavità amniotica in formazione
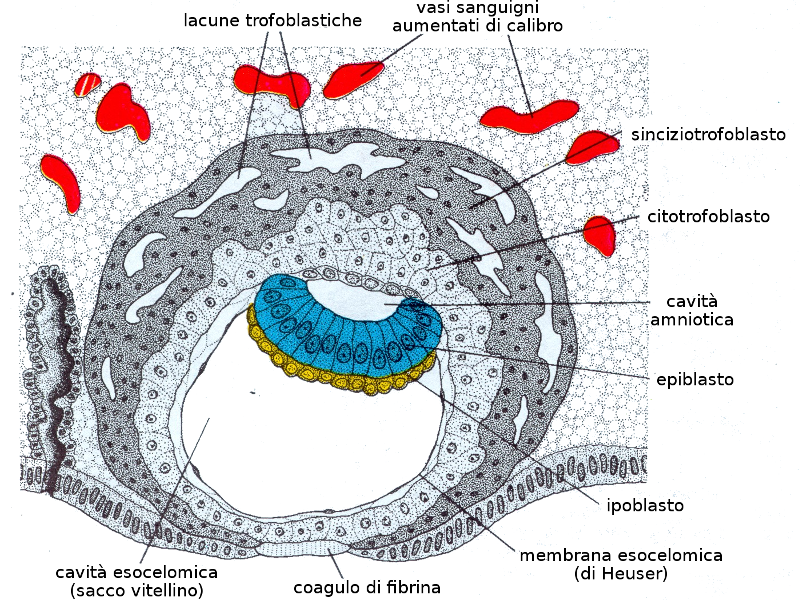
Il sinciziotrofoblasta produce Gonadotropina corionica, un ormone che impedisce la degenerazione del corpo luteo fino al quarto mese di gravidanza. Il corpo luteo dell'ovaio secerne il progesterone necessario per mantenere l'endometrio in condizione di poter mantenere la gravidanza. A partire dal quarto mese sono le cellule del trofoblasto che producono il progesterone. Qualsiasi trattamento che provoca l'inattivazione del corpo luteo prima del quarto mese, produrrà un aborto. A partire dalla seconda settimana di gravidanza si raggiungono livelli di gonadotropina corionica nel sangue sufficienti per essere misurati dai normali test di gravidanza.
Verso il nono giorno, si produce una membrana di cellule che vanno a rivestire la strato interno del citotrofoblasta, la cavità esocelomica, dando luogo al futuro sacco vitellino.
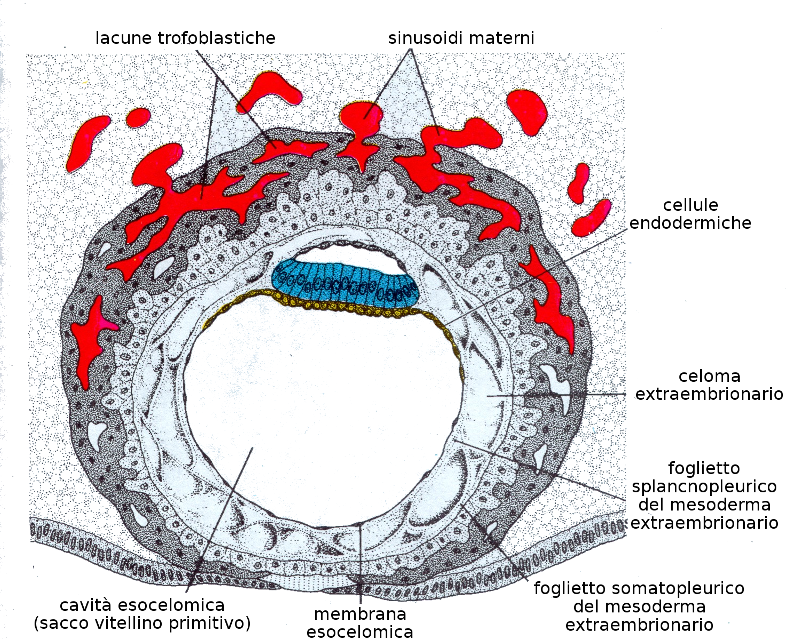
Verso l'undicesimo o dodicesimo giorno di sviluppo, la blastocisti è incluso al completo nello stroma endometriale, e l'epitelio superficiale copre quasi completamente la parete uterina. La blastocisti si approfondisce nella parete uterina. Inoltre, le cellule del sinciziotrofoblasta cominciano a infiltrarsi di più nella mucosa uterina e grazie a un intenso processo di angiogenesi il sangue materno comincia a fluire nel sistema trofoblastico: è l'inizio della circolazione uteroplacentare. La crescita del disco germinativo bilaminare è relativamente lento a paragone con il trofoblasto.
Verso il tredicesimo giorno di sviluppo l'endometrio è completamente cicatrizzato. Tuttavia a volte c'è una emorragia nel sito di impianto come conseguenza dell'aumento del flusso sanguigno nella circolazione uteroplacentare. Dato che questa emorragia si produce verso il ventottesimo giorno del ciclo mestruale, può confondersi con una mestruazione normale e causare equivoci nel calcolo del giorno del parto.
Riassunto
4. Gastrulazione e formazione della Notocorda
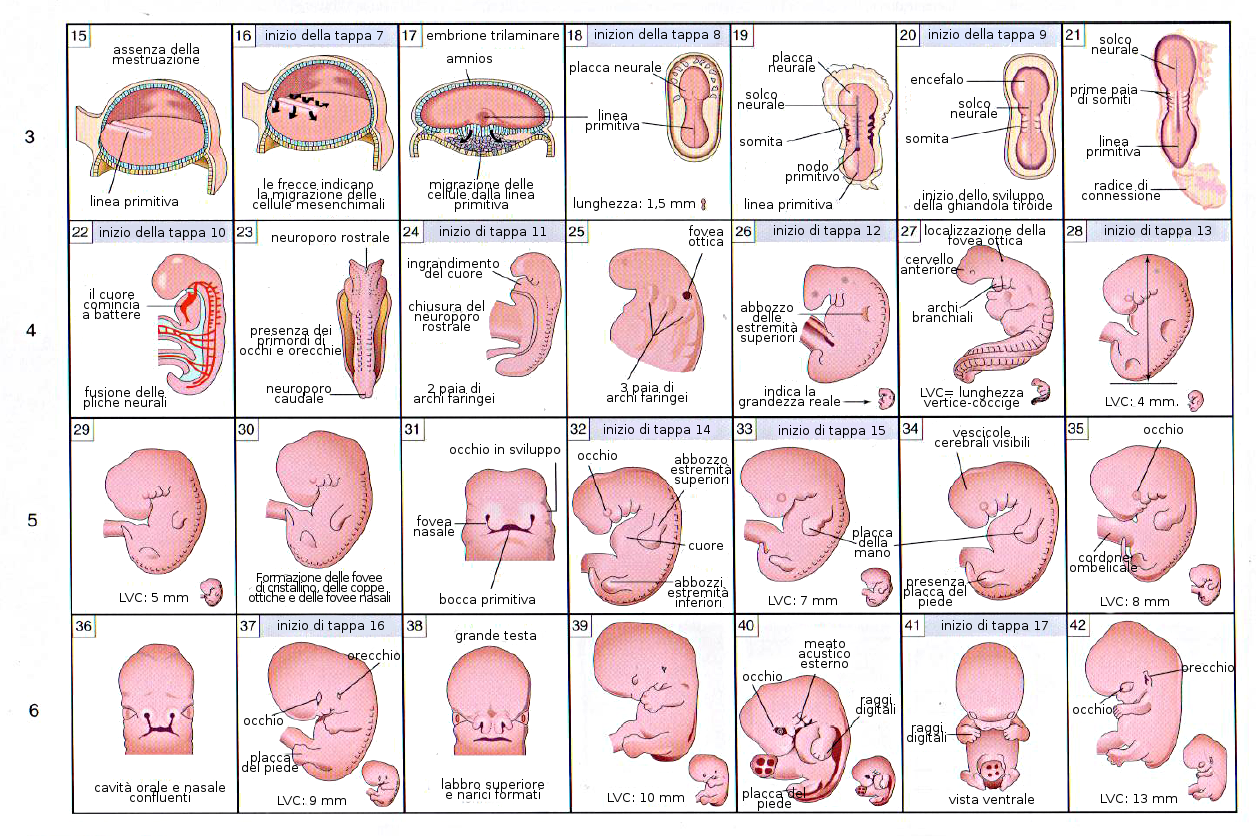
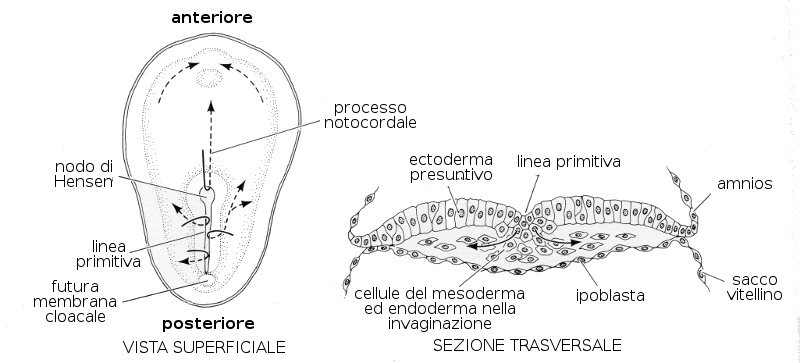
Il fenomeno più caratteristico che avviene durante la terza settimana di sviluppo è la Gastrulazione. È la tappa dello sviluppo embrionario nella quale grazie a movimenti cellulari si stabilizzano i tre foglietti germinativi a partire dall'epiblasta: l'ectoderma, l'endoderma e il mesoderma. Porta con se anche l'orientamento assiale dell'embrione. Questa tappa comincia con la comparsa della linea primitiva, che è un ingrossamento con un solco che compare nella zona dorsale media dell'epiblasta e che è già evidente a partire dal sedicesimo giorno. All'estremità cefalica contiene un rigonfiamento che prende il nome di nodulo primitivo o Nodulo di Hensen.
• Ectoderma embrionario: epidermide, SNC e SNP, retina dell'occhio, ...
• Endoderma embrionario: epiteli, ghiandole e organi acclusi (fegato, pancreas) dell'apparato digerente e vie respiratorie.
• Mesoderma embrionario: muscolatura liscia, connettivo e vasi degli organi, quasi tutto l'apparato cardio-vascolare, le cellule del sangue, il sistema muscolo-scheletrico, organi riproduttivi ed escretori.
A partire dalle cellule dell'epiblasta che migrano in direzione della linea primitiva si forma il Mesoderma e l'Endoderma intraembrionario. Questo processo di formazione del mesoderma ed endoderma si conosce come invaginazione, poiché le cellule distaccatesi dall'epiblasta si dislocano verso il basso rispetto a queste. Le cellule che rimangono nell'epiblasta costituiscono l'Ectoderma. Per tanto i tre foglietti embrionari si formano a partire dall'epiblasta.
Altro processo che avviene nell'epiblasta è la formazione della notocorda. La notocorda è un bastoncino più o meno rigido che si forma durante lo sviluppo embrionario e che caratterizza il gruppo di animali denominati Cordati. La notocorda costituisce l'asse scheletrico (scheletro assiale) primitivo e successivamente sarà sostituita dalla colonna vertebrale. A partire da alcune cellule della fossetta primitiva e, successivamente, da cellule dell'endoderma primitivo si va formando questo cordone.
5. La Neurulazione
La neurulazione è il processo di formazione del sistema nervoso centrale ed è il primo processo della organogenesi. Prima di spiegare il processo di neurulazione conviene aver chiari determinati concetti di embriogenesi come quelli di induzione o territorio competente.

5.1 La Induzione
L'induzione è un processo mediante il quale una regione embrionaria interagisce come una seconda regione, per far si che quest'ultimo tessuto si differenzi in una direzione che in altro modo non sarebbe seguito. Questo tessuto riceve il nome di territorio competente, vale a dire, la zona dell'embrione capace di rispondere in maniera specifica allo stimolo dell'induzione. Nel caso che seguirà, si vedrà che l'ectoderma dorsale è competente per l'induzione della notocorda.
Gli induttori sono molecole liberate dalle cellule di una determinata regione dell'embrione, ad esempio la activina o proteína FGF (Fattore di accrescimento di fibroblasti). La induzione è limitata nello spazio, poiché avviene solo nei tessuti adiacenti, e nel tempo poiché, ad esempio, a partire dalla gastrulazione va diminuendo la capacità di attivazione.
- induzione primaria: è il meccanismo induttivo per il quale si produce la struttura generale dell'embrione. Un esempio, nel caso dei vertebrati, è quello della notocorda.
- induzione secondaria: è il processo mediante il quale i tessuti indotti, determinati dalla notocorda e, infine differenziati, sono capaci a loro volta di indurre cambi negli altri tessuti adiacenti. Un esempio è l'induzione della formazione dell'occhio nella organogenesi oculare.
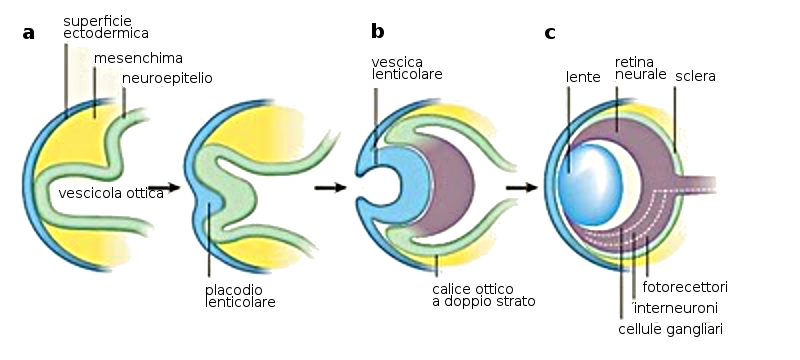
5.2 La Neurulazione
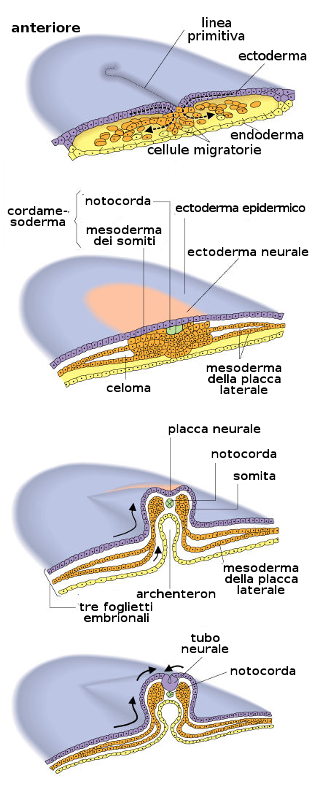
La neurulazione è il processo di formazione del sistema nervoso centrale grazie alla induzione della notocorda sopra l'ectoderma che è il territorio competente.
3. Neuroporo anteriore; 4. Neuroporo posteriore; 5. Cresta Neurale; 6. Tubo neurale; 7. Ectoderma; 9. Notocorda; 10. Solco neurale.
Per effetto della induzione della notocorda, l'ectoderma che si trova in cima a questa si ingrandisce formando la placca neurale. Poco prima della fine della terza settimana (giorno 18) i bordi laterali della placca neurale si elevano e formano le pliche neurali.
La porzione sommersa forma il solco neurale. Poco a poco le pliche neurali si dispongono attorno alla linea mediana, dove si fondono. Nella misura in cui le pliche neurali si elevano e si fondono, alcune cellule si staccano a formare una popolazione che costituisce la cresta neurale. Questa cresta penetra lateralmente nel mesoderma sottostante formando i gangli rachidei e i nervi, le cellule di Schwann che formeranno la guaina di mielina degli assoni dei nervi, le meningi, per la precisione la pia madre e la aracnoide, la midollare delle ghiandole surrenali.
Questo processo di neurulazione è asimmetrico, poiché comincia nella regione del futuro collo (quarto somita) e avanza in direzione cefalica e caudale, andando davanti alla prima. Come conseguenza della invaginazione dell'ectoderma si forma un tubo longitudinale vuoto aperto agli estremi caudale e cefalico. Questi orifizi ricevono il nome di neuropori caudale e craniale. Il neuroporo craniale si chiude approssimativamente nel giorno 25 (periodo da 18 a 20 somiti), mentre il neuroporo caudale lo fa nel giorno 27 (periodo dei 25 somiti).
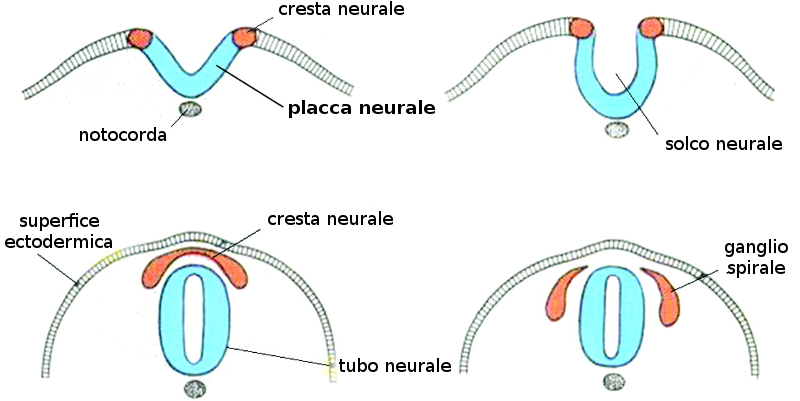
Il processo di neurulazione è completato e il sistema nervoso centrale è rappresentato da una struttura tubolare chiusa con una porzione caudale stretta, il midollo spinale, e una porzione cefalica molto più grande caratterizzata da varie dilatazioni, le vescicole cerebrali. Queste vescicole andranno segmentandosi e suddividendosi in direzione dell'asse antero-posteriore grazie all'azione dei geni HOX, specialmente in quello che concerne il rombencefalo.
In generale controllano lo sviluppo morfologico del corpo per quanto riguarda i segmenti lungo l'asse antero-posteriore.
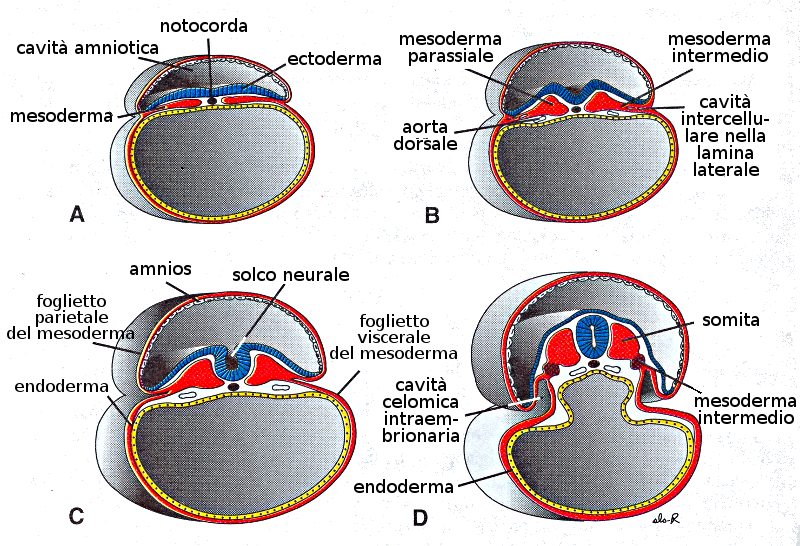
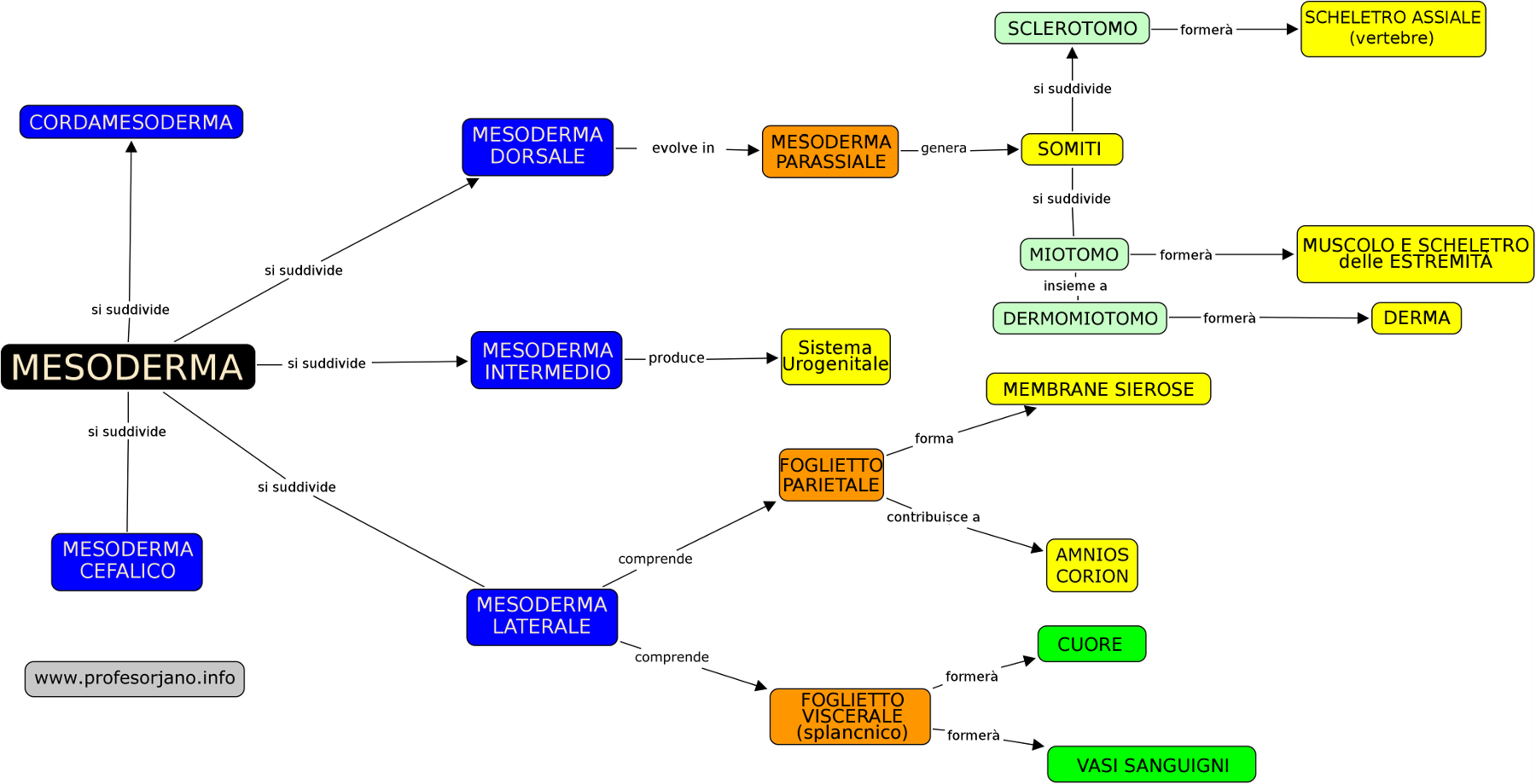
6. L'evoluzione del Mesoderma
All'inizio le cellule del foglietto germinativo mesodermico forma una lamina tenue di tessuto lasso a ogni lato della linea media. Tuttavia verso il diciassettesimo giorno (e in forma sincrona con l'evoluzione prima esposta dell'ectoderma) cominciano una serie di proliferazioni e ingrossamenti che daranno distinti tipi di mesoderma:
- Cordamesoderma: è il mesoderma che origina la notocorda.
- Mesoderma dorsale: darà il mesoderma parassiale. Questa è una zona ingrossata nella zona media del mesoderma.
- Mesoderma laterale: nella regione laterale, il mesoderma continua ad essere sottile. Questo si divide in due regioni:
- Foglietto somatico o parietale: ricopre l'amnios.
- Foglietto splancnico o viscerale: ricopre il sacco vitellino. Tra queste due si forma una nuova cavità: la cavità celomica intraembrionaria.
- Mesoderma intermedio: è quello che unisce il mesoderma parassiale con il laterale.
- Mesoderma cefalico
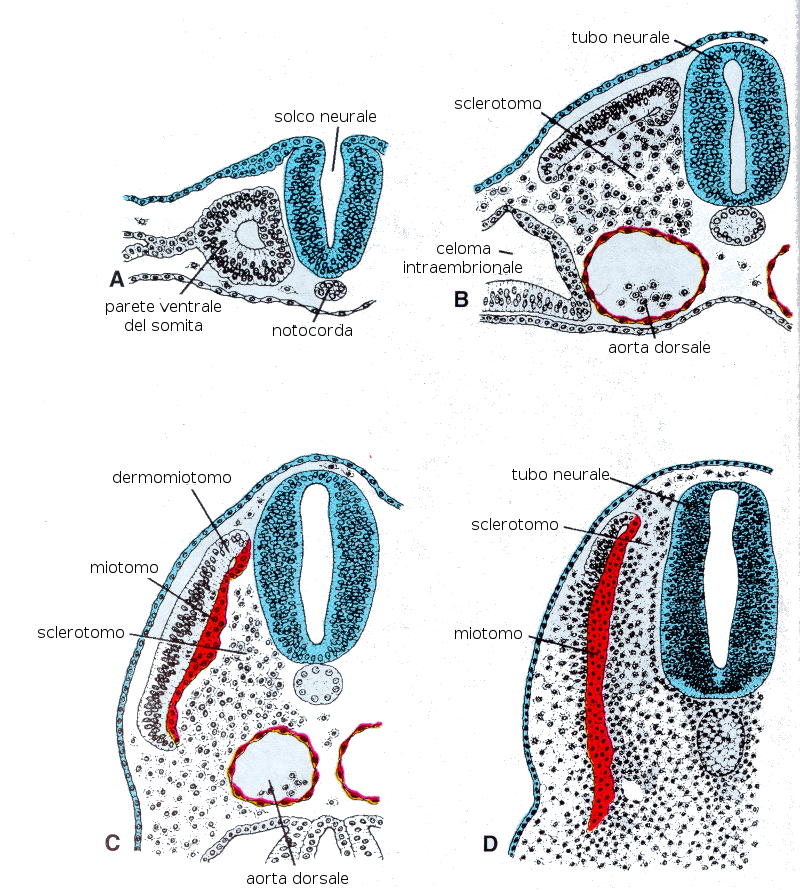
6.1 La formazione dei somiti
I somiti sono ognuno dei segmenti che si producono nel mesoderma parassiale e che cominciano a prodursi attorno alla regione cefalica-cervicale verso l'inizio della terza settimana (a partire dal giorno 20-21 appare il primo). Questo processo comincia grazie alla espressione del gene c-hairy nelle cellule del mesoderma parassiale. A partire da questo punto si formano nuovi somiti in direzione cefalocaudale, più o meno tre paia al giorno, fino a che alla fine della quinta settimana si trovano da 42 a 44 paia. Si distribuiscono in: 4 occipitali, 8 cervicali, 12 toracici, 5 lombari, 5 sacrali e 8-10 coccigei. Successivamente scompaiono alcuni di questi somiti.
Differenziazione del somita
A partire dalla quarta settimana alcune cellule dei somiti contornano la notocorda. La notocorda emette segnali chimici che attivano i geni Pax-1 e Pax-9 della metà ventrale del somita, il che provoca la differenziazione di alcune cellule. Queste cellule ricevono insieme il nome di sclerotomo e formeranno un tessuto lasso chiamato mesenchima. Contorneranno anche il midollo spinale e formeranno la colonna vertebrale. La notocorda andrà scomparendo.
Il resto del somita si differenzia in due strati:- Dermomiotomo: strato più esterno che darà il derma della pelle e produrrà l'altro strato del resto del somita:
- Miotomo: che produrrà lo strato muscolare del corpo che corrisponda alla sua localizzazione.
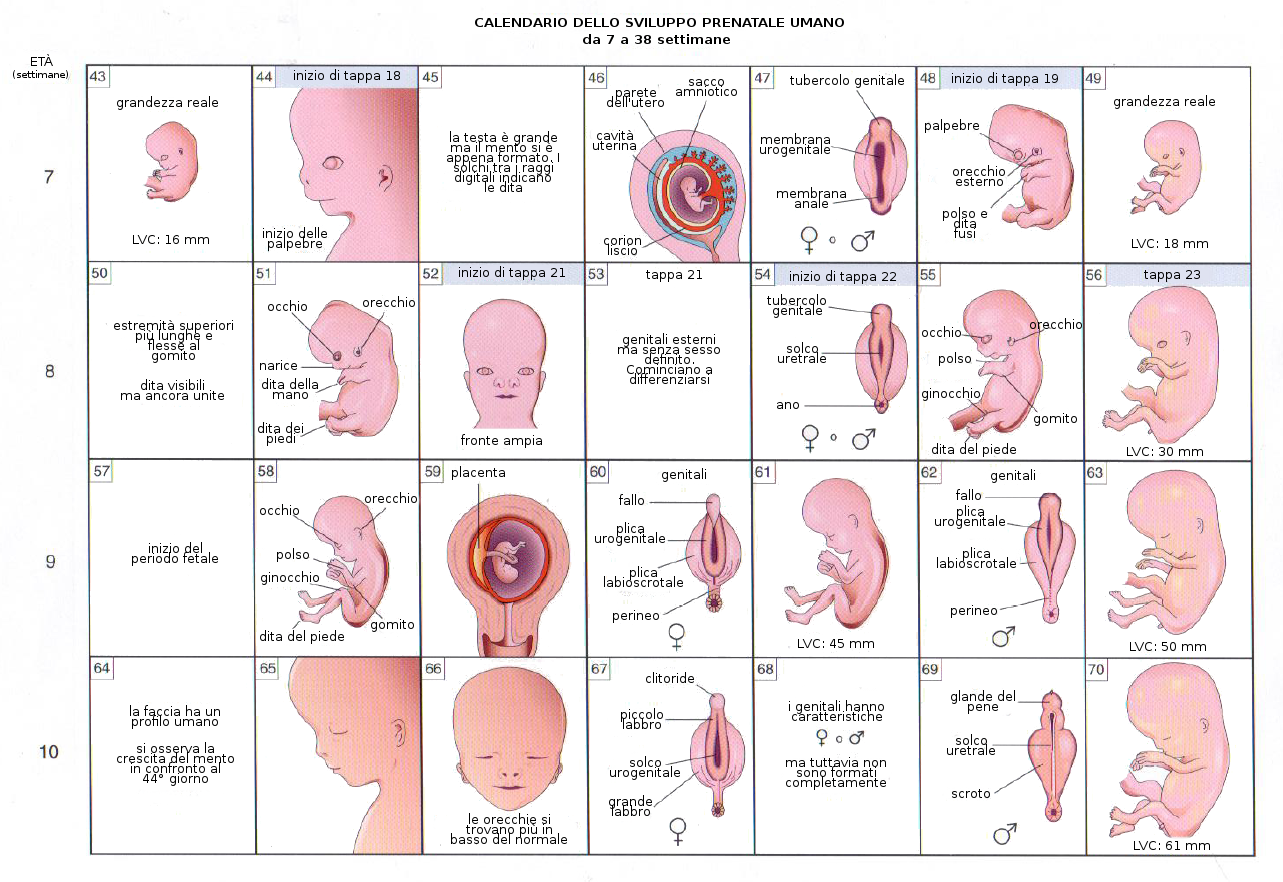
Il Mesoderma Intermedio congiunge temporaneamente il mesoderma parassiale con la lamina laterale. Si differenza in modo distinto rispetto a quello dei somiti e a partire dai cordoni di cellule del mesoderma intermedio se producono i reni e le gonadi. La differenziazione dell'apparato genitale comincia nella regione cefalica con la formazione del pronefrone o rene primitivo a partire del mesoderma intermedio di questa regione.
Ugualmente si differenzia in ogni lato dell'embrione il dotto pronefrico attorno al quale si organizzerà definitivamente l'apparato urogenitale.
Il Mesoderma Laterale come come già scritto, presenta due foglietti differenti, parietale e viscerale, generandosi all'interno di entrambi il celoma intraembrionario. Il foglietto parietale insieme all'ectoderma che lo ricopre, formerà le pareti corporee. Il mesoderma viscerale insieme all'endoderma embrionario formerà le pareti dell'intestino. Questo mesoderma (viscerale) formerà anche le membrane sierose: il pericardio, la pleura e il peritoneo. Formerà anche il mesentere.
A partire dalla terza settimana e nel foglietto viscerale che forma parte del sacco vitellino si differenziano gli angioblasti o cellule che produrranno i vasi sanguigni di molti vísceri come il pancreas e il fegato.
7. L'evoluzione dell'Ectoderma
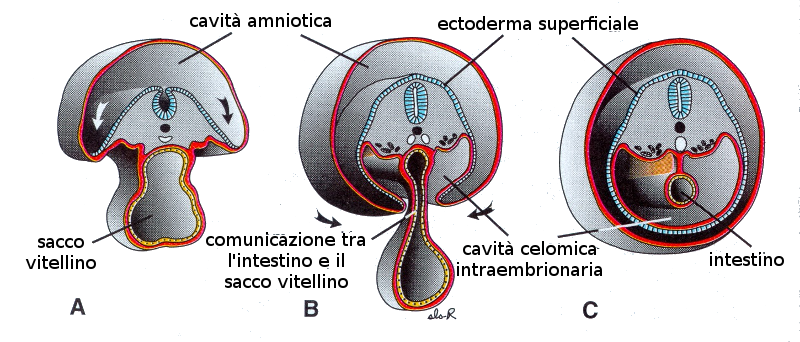
Il tratto gastrointestinale è il principale derivato del ripiegamento dell'endoderma. Si tratta di un processo passivo, vale a dire, come conseguenza di altro movimenti che si producono nell'embrione e non per attività propria. Questi movimenti sono essenzialmente due:- Ripiegamento cefalo-caudale, che si produce come conseguenza della neurulazione.
- Ripiegamento trasversale: come conseguenza della formazione dei somiti.
Questi ripiegamenti provocano l'incorporazione di parte del sacco vitellino rivestito di endoderma nella cavità corporea generandosi così il tratto digestivo. Il tubo digestivo rimane connesso con il sacco vitellino attraverso un condotto che si restringe man mano che trascorrono i giorni: è il dotto vitellino o dotto onfalomesenterico. Questo condotto darà origine al cordone ombelicale.
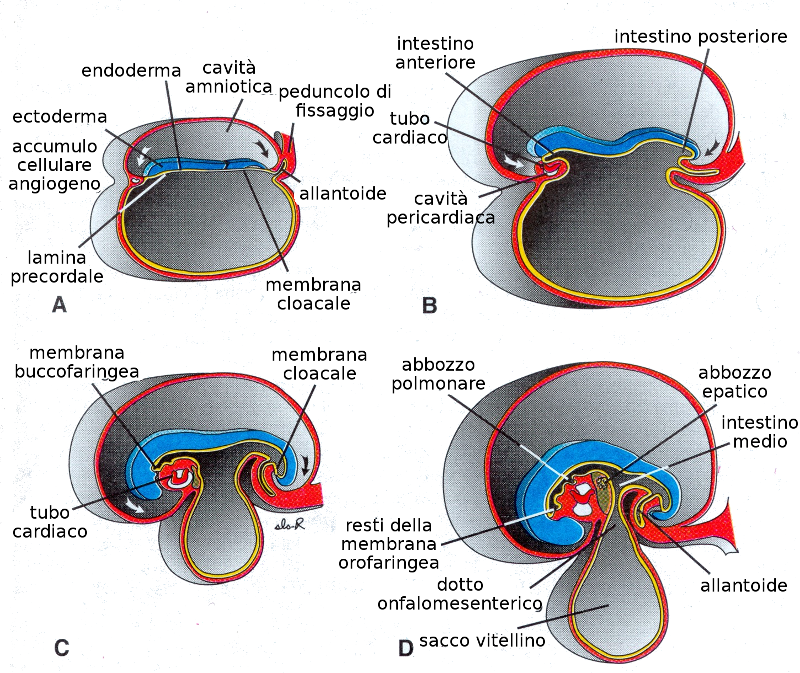
Il ripiegamento è più pronunciato nella zona della testa e della coda costituendo le cosiddette curve cefaliche e caudale. Il piegamento della regione caudale non è tanto pronunciato come quello della regione cefalica, ma una porzione di intestino posteriore si introduce nel mesoderma creando una borsa, l'allantoide, che nell'essere umano non ha una funzione separata, sebbene sembri che contribuisca all'interscambio di gas e alla eliminazione degli scarti embrionari.
Una membrana chiamata membrana orofaringea ricopre l'estremità cefalica del tubo digestivo. A partire dalla terza settimana si rompe e così si stabilisce una comunicazione tra l'intestino primitivo anteriore e l'amnios. Nell'estremità dell'intestino posteriore si trova la membrana cloacale. L'intestino primitivo medio continua con il sacco vitellino.

8. Testa e collo. Archi faringei
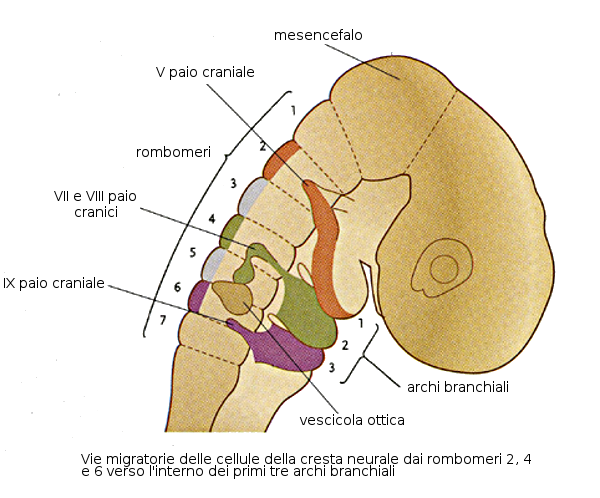
A partire dal mesoderma parassiale si forma la base del cranio e parte dell'occipite, oltre ai muscoli della testa. il mesoderma della lamina laterale forma le cartilagini laringee e le cellule della cresta neurale formeranno il resto degli elementi della faccia, archi faringei, meningi e neuroni sensitivi.
La caratteristica più tipica dello sviluppo della testa e del collo è la formazione degli archi branchiali o faringei a partire dalla 4a-5a settimana. Questi archi cominciano come delle piccole sacche di mesenchima rivestite di endoderma nella regione faringea. Inoltre esiste un diverticolo impari nella zona media chiamato primordio tiroideo. Le prime sacchette faringee contribuiranno alla formazione dell'orecchio. L'ectoderma di ognuna di queste regioni contiene un paio di fenditure laterali denominate fenditure branchiali.
Pertanto gli archi faringei sono queste masse di mesenchima alternate da queste fenditure. La formazione di questi archi branchiali inizia con l'espressione, in questa zona, dei geni HOX B.
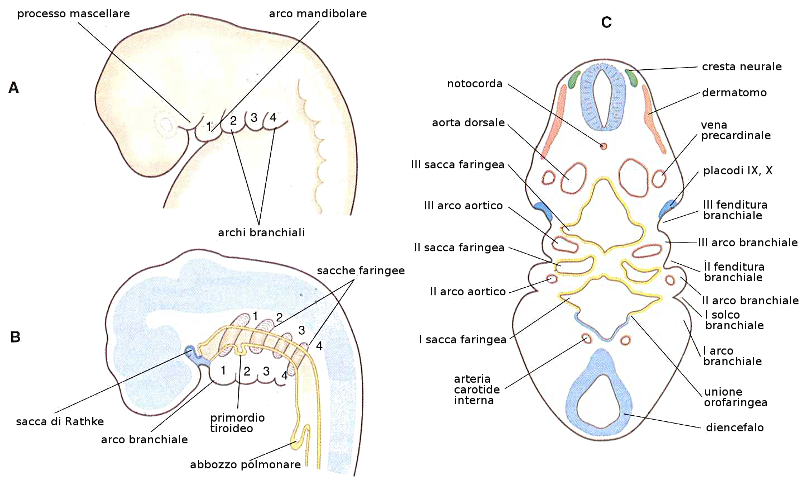
- Primo arco: Contiene nella regione dorsale il processo mascellare che darà origine al mascellare superiore ed altre ossa.
Nella zona ventrale se trova il processo mandibolare che contiene la cartilagine di Meckel a partire dalla quale si formerà l'incudine e il martello dell'orecchio medio. Questo processo mandibolare genererà anche la mandibola.
Dal primo arco evolveranno i muscoli della masticazione e il V paio di nervi cranici, il trigemino. - Secondo arco: È l'arco ioideo e da luogo all'ossicino della staffa e al VII paio di nervi cranici, i nervi cranio-facciali.
- Terzo arco: Origina strutture relazionate con l'osso ioide e il IX paio di nervi cranici, il glossofaringeo.
- Quarto arco: Produrrà alcuni muscoli e cartilagini del faringe. L'innervazione di questi muscoli dipende dal nervo vago, o X paio. Da parte delle sacche faringee del III e IV arco deriveranno le paratiroidi.
9. Avvolgimenti Embrionari
Il trofoblasto è una parte importante degli avvolgimenti che coprono e connettono il feto alla madre. A partire dal secondo mese di gravidanza, il trofoblasto si caratterizza per abbondanza di villosità secondarie e terziarie che gli danno un aspetto abbastanza ramificato.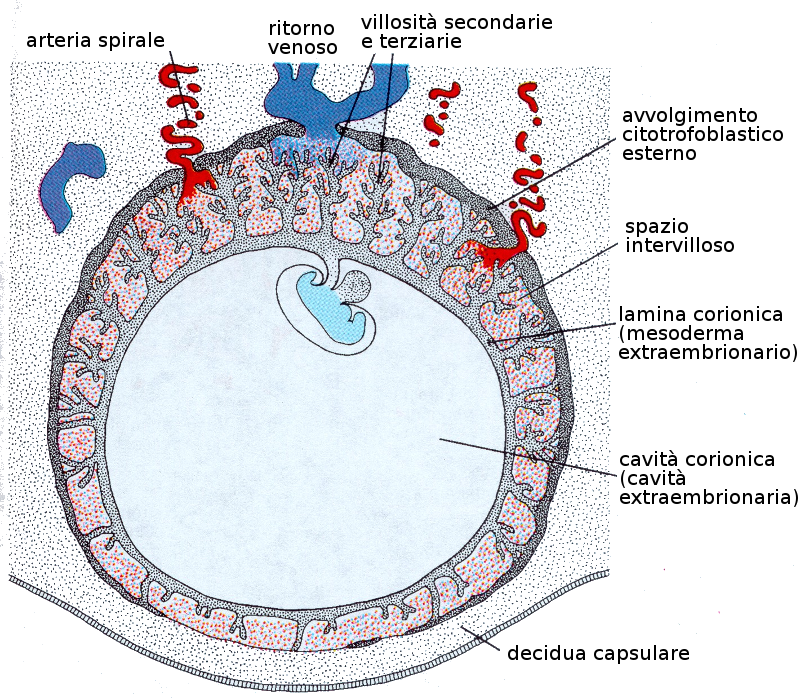
La crescita del trofoblasto è asimmetrico. Le villosità del polo embrionario continuano a crescere ed espandersi dando luogo al corion frondoso. Quelle del polo abembrionario o vegetativo degenerano e lasciano il corion tenue.
La zona del corion frondoso è interdigitato con uno strato compatto di cellule dell'endometrio materno che costituiscono la decidua che è lo strato dell'utero che si distaccherà durante il parto. Il corion frondoso insieme alla decidua forma la placenta che, pertanto, è una struttura formata tanto dal feto come dalla madre.
9.1 Struttura della placenta
Come si è già detto, la placenta possiede due componenti:- Una porzione fetale formata dal corion frondoso.
- Una porzione materna costituita dalla decidua basale.
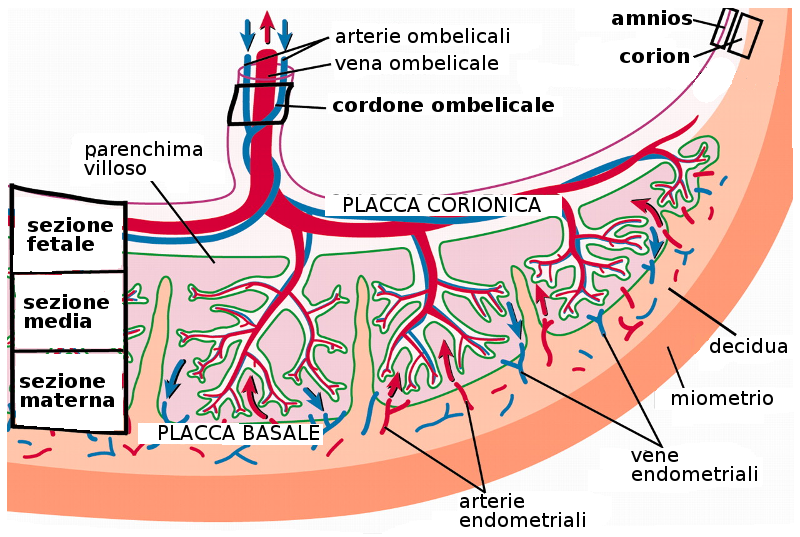
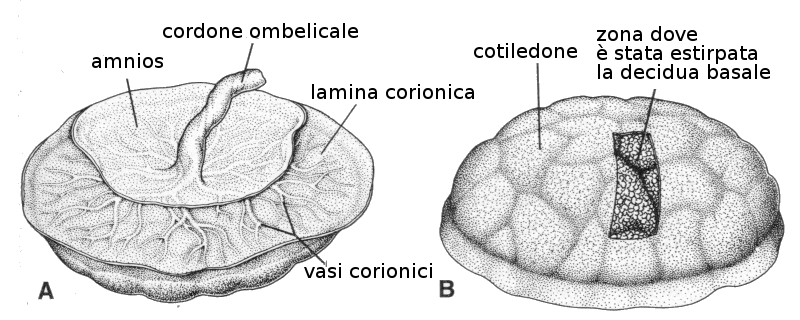
Al termine della gravidanza la placenta è una struttura con forma di disco di 15-25 cm di diametro e 3 cm di spessore. Pesa 500-600 gr. Nel momento del parto si distacca dalla parete uterina e circa 30 minuti dopo è espulsa dalla cavità uterina.
All'osservazione di questa placenta espulsa si vede che dalla parte materna ci sono 15-20 zone prominenti: sono i cotiledoni separati da trabecole deciduali. La superficie fetale è ricoperta completamente dal corion con arterie e vene di largo diametro, che costituiscono i vasi corionici. Questi vasi convergono nel cordone ombelicale che contiene due arterie e una vena. Questo corion è ricoperto a sua volta dall'amnios.
9.2 Circolazione della placenta
I cotiledoni della placenta ricevono sangue attraverso le arterie spirali. Questo sangue è raccolto dalle vene endometriali.
Il sangue del feto e della madre non entrano in contatto per via degli strati che li separano, come l'endotelio dei vasi sanguigni del feto, il tessuto connettivo delle villosità e il citotrofoblasto. Queste strutture costituiscono la barriera placentaria che è molto lontano dall'essere una barriera impermeabile, poiché permette il passaggio di molte sostanze.
9.3 Funzioni della placenta
- Interscambio di gas, il diossido di carbonio e l'ossigeno per diffusione semplice. Approssimativamente, il feto estrae da 20 a 30 ml di ossigeno al minuto dalla circolazione materna. Il feto è molto sensibile alla interruzione nell'apporto di ossigeno.
- Interscambio di nutrienti, come aminoacidi, glicidi, acidi grassi liberi o vitamine. Assorbe anche i prodotti di scarto del feto.
- Trasmissione di anticorpi materni. Sono captati dal sinciziotrofoblasto e trasportati poi verso i capillari fetali. Questi anticorpi sono immunoglobuline d tipo G.
- Produzione di ormoni. A partire dal quarto mese, il progesterone che mantiene la gravidanza è prodotto dalla placenta che potrebbe sostituire quello secreto dal corpo luteo. Produce anche estradiolo e somatomammotrofina che determina la priorità del feto sulla madre nella captazione del glucosio circolante. Inoltre stimola lo sviluppo delle mammelle per la produzione del latte.
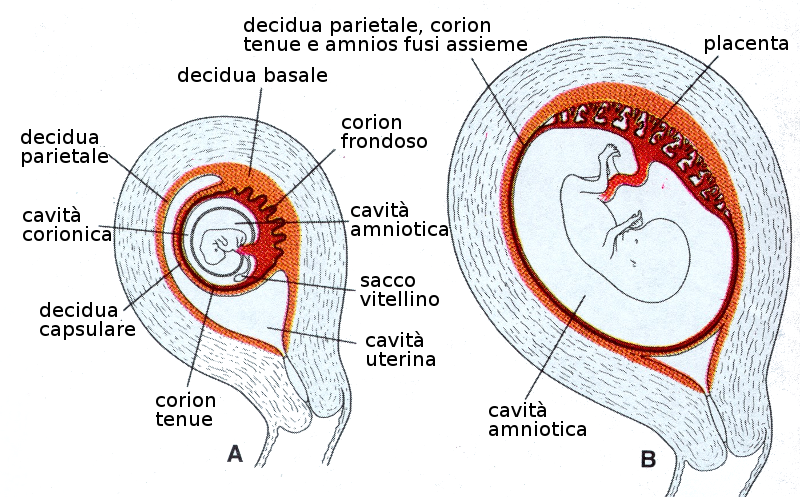
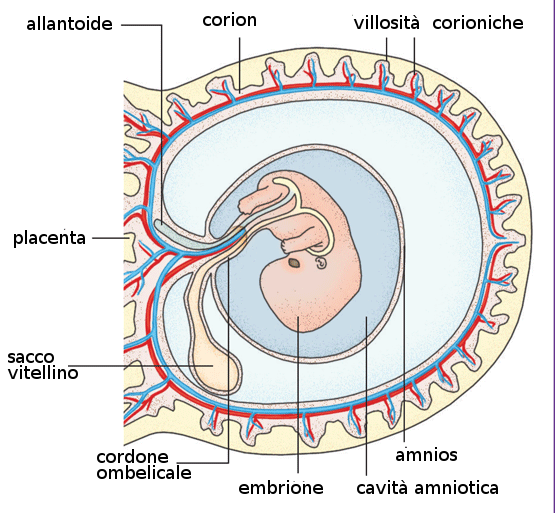
9.4 Amnios e Cordone ombelicale
Le pliche dell'amnios nella regione ventrale dell'embrione insieme all'ectoderma embrionario formano l'anello ombelicale primitivo. Da questa struttura e a partire dalla quinta settimana comincia a formarsi il cordone ombelicale propriamente detto che contiene:- Un peduncolo di fissazione al corion.
- Una porzione del sacco vitellino insieme con anse intestinali.
- Vasi sanguígni
A partire dal terzo mese le anse intestinali ritornano nel corpo del feto e l'allantoide e il sacco vitellino collassano fino a scomparire. In questo modo il cordone ombelicale rimane formato dai vasi ombelicali e un tessuto che lo contorna, chiamato gelatina di Wharton.
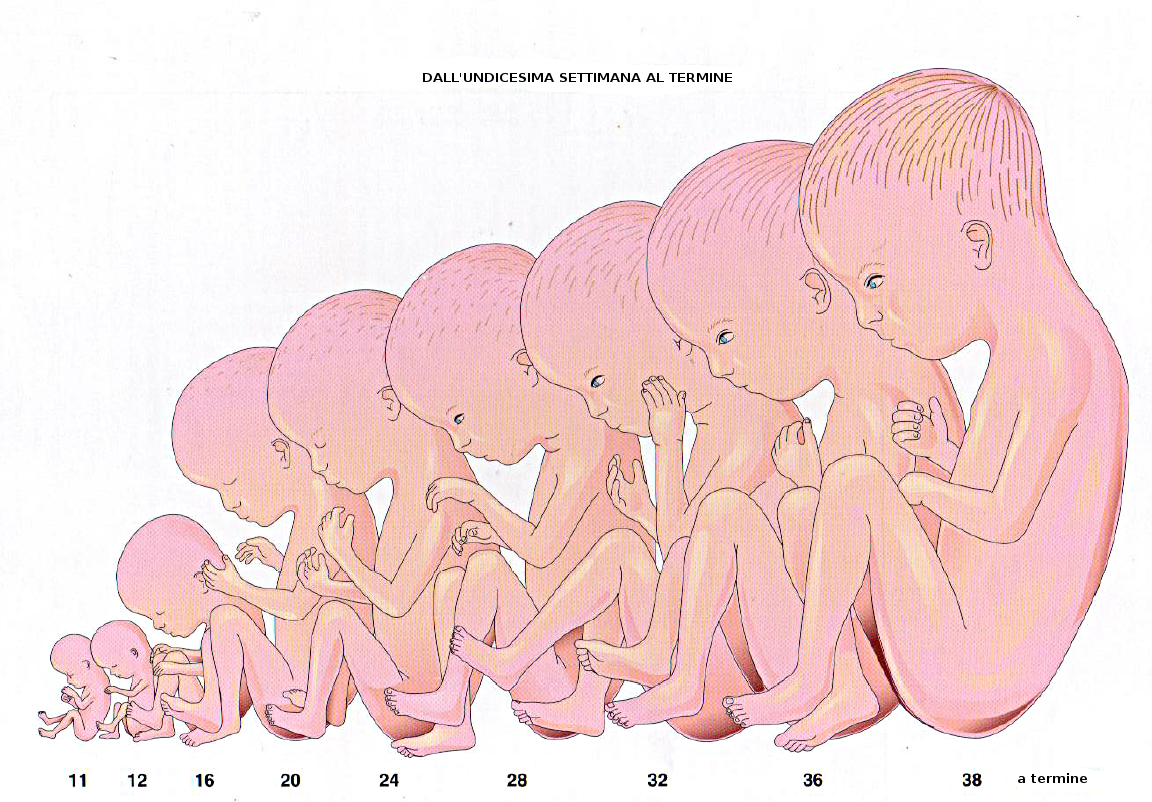
La cavità amniotica contiene un liquido acquoso e cristallino che proviene delle cellule proprie dell'amnios e anche, soprattutto al principio, dal sangue materno. Nella decima settimana di gravidanza la quantità di liquido amniotico è di circa 30 mL, 350 alla ventesima settimana e tra 800 e 1000 mL alla 37ma settimana. Questo liquido è sostituito ogni tre ore.
- Ammortizzare gli scossoni
- Impedisce che l'amnios si attacchi all'embrione.
- Facilita i movimenti fetali.